【国立科学博物館】あえて「臭く」進化した花たちのニオイを生み出す仕組みを解明-虫を呼ぶために複数の植物で収斂進化していた!-
国立科学博物館(館長:篠田謙一)の研究主幹 奥山雄大(植物研究部・筑波実験植物園/東京大学大学院理学系研究科准教授兼任)は、国立遺伝学研究所、昭和医科大学、長野県環境保全研究所、宮崎大学、東北大学、情報・システム研究機構ライフサイエンス統合データベースセンター、龍谷大学、慶應義塾大学との共同研究により、腐った肉のような臭いニオイで昆虫をだまして花粉を運ばせる(腐肉擬態)花が、臭いニオイの成分「ジメチルジスルフィド(※1)」を生み出すメカニズムを解明し、またその機能を獲得する進化がわずかなアミノ酸置換でもたらされることを実験的に示すことに成功しました。さらにそのメカニズムを担う酵素がカンアオイ属(※2)、ヒサカキ属、ザゼンソウ属という全く異なる植物で独立に進化、獲得されていることを発見しました。これは、花による腐肉擬態というユニークな現象が、どのような成り立ちで進化しうるのかを明快に説明できた類い稀な成果と言えます。本研究成果は、2025年5月8日刊行のScience誌に掲載(発表)されました。
研究のポイント
-
カンアオイ属を対象に、花の臭いニオイ成分「ジメチルジスルフィド」を含む種と含まない種の間で比較研究を行い、臭いニオイの生合成に関与する複数の遺伝子を特定。
-
特定した遺伝子の一つがジメチルジスルフィドを生合成する新発見の酵素の遺伝子であることを解明し、これをジスルフィドシンターゼ(DSS)と命名。
-
DSSの機能は陸上植物が共通して保有する祖先的な酵素メタンチオールオキシダーゼからわずかなアミノ酸配列の変化で獲得されることを解明。
-
DSS はカンアオイ属だけでなく、全く異なる植物のグループであるヒサカキ属、ザゼンソウ属でも 独立に進化し、全く同じプロセスを経て同じ機能を持つ酵素を獲得したことを発見。
-
植物園が戦略的に収集して維持しているリビングコレクション(生植物)が、未知の生命現象を解き明かすために決定的に重要な役割を果たしうることを示した。
研究成果の詳細について
【発表論文】
表 題:Convergent acquisition of disulfide-forming enzymes in malodorous flowers.
著 者:Okuyama, Y., Fukushima, K., Kakishima, S., Valchanova, A. K., Takano, K. T.,
Ito-Inaba, Y., Nakazato, T., Nagano, A. J.
掲載紙:Science
(アメリカ科学振興協会発行の科学雑誌、2025年5月8日付)
(URL):https://www.science.org/doi/10.1126/science.adu8988
DOI:10.1126/science.adu8988
※本研究は、科研費(JP19H03292, JP20H02917)、科学技術振興機構(JST)戦略的創造研究推進事業 さきがけ(JPMJPR21D3)の支援を受けています。
【研究の背景】
花といえば、美しい色や良い香りで私たちを楽しませてくれる存在と一般に捉えられがちですが、実際にはそのイメージにそぐわないさまざまな姿かたち、性質を持つ花が存在します。花が持つこの顕著な多様性は、花がそれぞれどのような方法で昆虫などの花粉を運ぶ動物を招き、受粉を助けてもらっているかという生態と密接に結びついています。中でも奇妙な花として特筆に値するのが腐った肉や糞(ふん)のようなにおいでハエなどの昆虫を騙し、花粉を運ばせる花(腐肉擬態花)で、このような性質を持つものには世界最大の花として有名なラフレシアやショクダイオオコンニャクが含まれます(図1)。腐肉擬態花は、あまりに特徴的な花の姿とその強烈なにおいのため、古くは18世紀末から人類に認識されていた典型的な生物擬態の一例ですが、どのようにしてこのような特異な花が進化したかについてはこれまでほとんど分かっていませんでした。
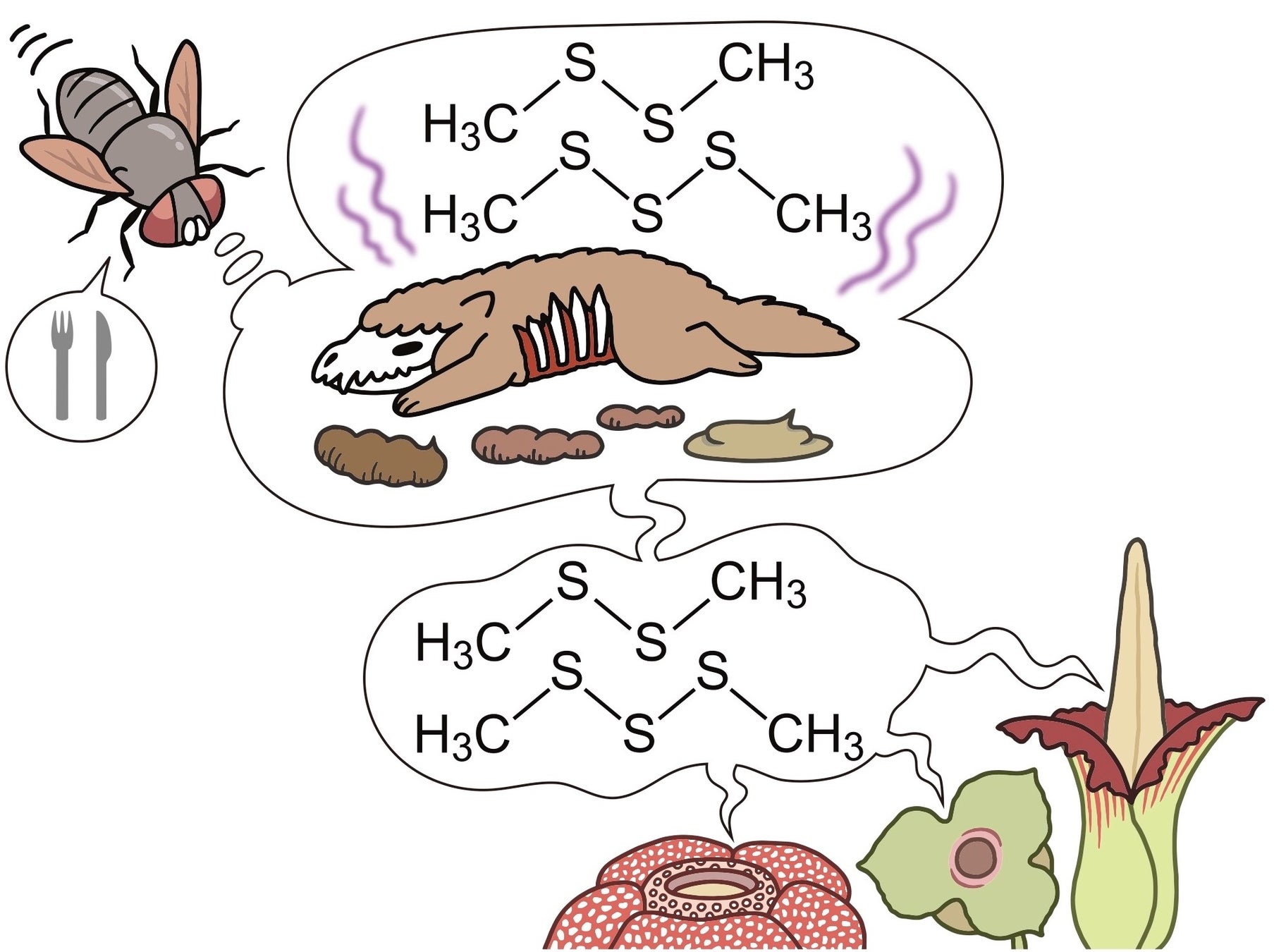
図1:腐肉擬態花の模式図。花からは腐った肉や肉食動物の糞から出るのと同じジメチルジスルフィド、ジメチルトリスルフィドといったにおい成分を放ち、花粉を運ぶハエなどの昆虫を誘き寄せる。
イラスト:いずもり・よう
例えば腐肉擬態花を特徴づける典型的な性質のひとつに、腐った肉などが放つ硫黄を含むにおい物質「ジメチルジスルフィド」を発するというものがあります。腐肉擬態花が進化する過程ではこの性質の獲得が必要不可欠ですが、そもそも腐肉擬態花がどのような仕組みでこのにおい物質を作り出しているのかについても未解明でした。私たちは、日本で50種あまりが自生し顕著な多様化を遂げた植物のグループであるカンアオイの仲間の中に、典型的な腐肉擬態花の性質を持ち花のにおいにジメチルジスルフィドを含む種と、そうではない(ジメチルジスルフィドを含まない)種が存在し、その進化の歴史において臭いにおいの花が繰り返し生じてきたことを見出しました。そこでこれらの種間で比較研究を行うことで花が臭いにおいを生み出す仕組みを解明できるのではないかと着想しました。
【研究の内容】

図2:本研究で明らかにした「臭い花」のにおい成分「ジメチルジスルフィド(DMDS)」の生合成経路。これまで、DMDSはメタンチオールが酵素反応を経ずに酸化して生じると考えられていたが、この反応を担う酵素ジスルフィドシンターゼ(DSS)を発見した。MGL: メチオニンガンマリアーゼ。MTOX: メタンチオールオキシダーゼ。
1.花が生合成するジメチルジスルフィドは、硫黄を含むアミノ酸の一つであるメチオニンに由来すると考えられてきました(図2)が、これも実証されてはいませんでした。そこでカンアオイの花にメチオニンの水溶液を与える実験を行い、このメチオニンがジメチルジスルフィドに変換されて花から放出されるかを調べました。この際与えたメチオニンは、自然界にはわずかしか存在しない炭素の安定同位体である炭素13で標識したメチオニンで、それを原料として生じるジメチルジスルフィドは本来の分子量である94のものではなく、分子量95や96のものとなるはずです。実際に質量分析計を用いた分析の結果、花に与えたメチオニンは確かにジメチルジスルフィドに変換されていることが確認されました(図3)。また花をすり潰して得た粗抽出液にも、メチオニンからジメチルジスルフィドを生合成する強い酵素活性が存在することも確認できました。さらにこの酵素活性は花の香りにジメチルジスルフィドを含む種にだけ顕著でした。このことは、花に含まれる酵素活性の違いが、花の香りにジメチルジスルフィドを含むか含まないかの違いの原因になっていることを示唆しています。

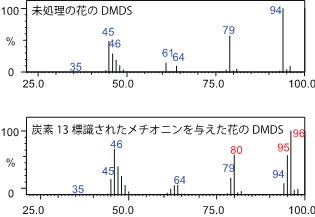
図3:カンアオイの花にメチオニン水溶液を与える実験の様子(左)と、花から放出されるジメチルジスルフィド(DMDS)のマススペクトル。(右)
右下段のマススペクトルには未処理の花にはほとんどみられない80, 95, 96といった質量数のシグナル(赤数字)が存在することから、花に吸わせたメチオニン(炭素13標識されたもの)がジメチルジスルフィド(DMDS)に変換されたことが分かる。
2.そこで次に、筑波実験植物園で栽培保有するカンアオイのリビングコレクション(生植物)から、花の香りにジメチルジスルフィドを含む種と含まない種を系統樹全体から均等に選んだ30系統(26種からなる)の花を用いて、系統関係を考慮に入れた全遺伝子発現プロファイル(トランスクリプトーム(※3))の比較解析を行いました。これにより花から放出されるジメチルジスルフィドの量と遺伝子発現量が強く相関する遺伝子を絞り込んだ結果、2つの遺伝子MGL(メチオニンガンマリアーゼ遺伝子)とSBP(セレン結合タンパク質遺伝子)が見出されました。これらはいずれも硫黄代謝と関係すると考えられる遺伝子です(図4)。ジメチルジスルフィドも硫黄を含むため、これらの遺伝子がジメチルジスルフィド生合成に関与している可能性を検討しました。


図4:カンアオイのリビングコレクション30系統の系統関係とそれぞれの花から放出されるDMDSの量およびMGL SBP遺伝子の発現量の関係。系統関係を補正した解析(PIC)により、両遺伝子の発現量とDMDSの量には強い相関があることが分かった。
この目的のためカンアオイの花から遺伝子クローニング(※4)して得た2つの遺伝子MGL及びSBPを大腸菌に組み込み、組換えタンパク質(※4)という形で大腸菌にその遺伝子産物(MGL及びSBP)を作らせ、これらの酵素としての機能を検証しました。この結果、MGLはメチオニンをメタンチオールという物質に変換する働きがあることが確認されました。
従来、このメタンチオールは自然に酸化しジメチルジスルフィドに変化するとされてきました(図2)。ところが実際にはMGL単独でメチオニンと反応させてもジメチルジスルフィドはほとんど生じず、通説にはそぐわない結果が得られました(図5)。これはメタンチオールがジメチルジスルフィドに変換される際にさらなる別の酵素反応が関わっている可能性が高いことを示しています。
そこで次にSBPの酵素としての働きを調べた結果、メタンチオールをジメチルジスルフィドに変換する働きが確認されました(図5)。このような働きを持つ酵素はこれまで知られておらず、本研究チームはこの新規酵素を「ジスルフィドシンターゼ(DSS)」と命名しました。
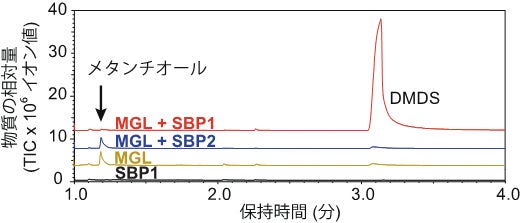
図5:カンアオイ属由来のMGLとSBPをメチオニンと反応させた生成物のガスクロマトグラム。メチオニンを基質とすると、MGL単独ではメタンチオールを生じる一方でDMDSはほとんど生じないが、SBP1はメタンチオールからDMDSを生成する。SBP2にはDMDSを生成する活性は見られず、別の実験からメタンチオールオキシダーゼ(MTOX)として働くことが分かった。
3.SBPと相同の酵素はヒトを含む動物やバクテリアにも存在し、その働きはDSSとは異なり、メタンチオールを硫化水素と過酸化水素に分解するメタンチオールオキシダーゼ(MTOX)であることが知られていました(図2)。しかし、植物での機能は未知でした。そこでカンアオイのSBPにも類似の働きがあるかどうかを調べました。詳細に調べた結果、カンアオイから遺伝子クローニングしたSBPにはそれぞれSBP1、SBP2、SBP3という3つのサブタイプがあり、SBP2にのみMTOXとしての働きがあることを確認しました。一方SBP1はDSSの働きを持つ一方でMTOXの働きは持たないことが分かりました。なおSBP3については機能は明らかにできませんでした。
さらにカンアオイのDSS(=SBP1)の起源を明らかにするため、コケ植物やシダ植物、裸子植物も含め幅広い陸上植物においてSBPの相同遺伝子をクローニングし、先の実験と同様大腸菌に組換えタンパク質を作らせることでその酵素としての働きを網羅的に調べました。その結果、MTOXとしての働きは陸上植物全体で高度に保存されていることが確かめられました(図6)。またこれら陸上植物のSBPとカンアオイ属のSBPの分子系統解析を行った結果、カンアオイ属においてはSBP2(=MTOX)が祖先的であり、そこからSBP1(=DSS)が派生して生じたことが明らかになりました。さらに、系統解析の結果推定復元されたカンアオイ属SBP1の祖先配列Yを人工的に再構築するとともに、その直近の祖先配列Xについても人工的に再構築し、その両者で酵素機能および配列を比較した結果、両者の分子の間ではたった5アミノ酸しか違いがないにも関わらず、MTOXからDSSへの機能転換を起こしていることを突き止めました(図6)。このことは、MTOXとDSSの間での機能転換がわずかなアミノ酸置換によって起こる比較的容易な形質進化であることを示唆しており、腐肉擬態花が被子植物の多くの系統で繰り返し進化できた理由である可能性があります。

図6:陸上植物におけるSBP遺伝子の分子系統樹とその遺伝子産物の酵素としての働き。この結果から、ピンク色の矢印の箇所で独立に遺伝子重複が起き、DSSとしての働きが獲得されていることが分かる。またカンアオイ属においては祖先配列XとYを人工的に遺伝子合成し、5アミノ酸の違いしかないその遺伝子産物の間で酵素機能の変化が起きたことも実証した。
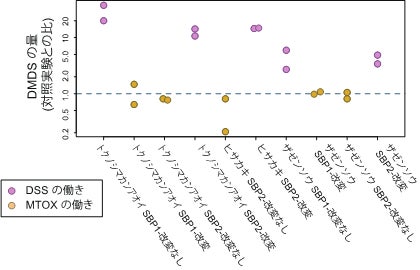
4.そこで次に、カンアオイ属の一部の種と同様、ジメチルジスルフィドを花から放出する「臭い花」の代表として、ショクダイオオコンニャク(サトイモ科)、ザゼンソウ(サトイモ科)、ヒサカキ(モッコク科)についても調査を行ったところ、少なくともザゼンソウとヒサカキでは、カンアオイ属と同様にMGLとSBPの両方の働きでジメチルジスルフィドが生合成されていることが明らかになりました。またザゼンソウ、ヒサカキにおいてはSBPにそれぞれ2つのサブタイプが存在し、それぞれDSSとMTOXの働きを持つことを突き止めました(図6)。そこで、遺伝子の分子系統樹を用いたタンパク質配列の収斂(れん)進化を検出する解析法を適用してカンアオイ属、ザゼンソウ属(ザゼンソウ)、ヒサカキ属(ヒサカキ)のSBPの間での分子収斂進化を探索した結果、4つのアミノ酸座位において、この3つの植物群で共通したアミノ酸置換が起きていることを突き止めました。この結果をもとに、この3つの植物群それぞれのDSSとMTOXをコードしているSBPサブタイプ間で、特定された座位のアミノ酸を入れ替えた配列を人工的に作成し、その遺伝子産物の酵素としての働きを調べた結果、このうち2ないし3のアミノ酸を入れ替えただけでDSSとMTOXの機能も入れ替わることが確認できました(図7、8)。これはSBP遺伝子の祖先的な機能であるMTOXをDSSへと変換する適応進化の原因はこの2ないし3座位におけるアミノ酸置換のみであり、しかも全く異なる3つの植物の系統で独自に、同じメカニズムによってDSSが獲得される進化が生じていることを示しています。つまり、「臭い花」への進化の道筋は限られており、独自に臭い花を進化させた3つの植物群は、全く同じプロセスを経て全く同じ機能を持つ酵素を獲得したということになります。本研究が明らかにしたのは顕著な分子収斂進化による新規機能獲得の明白な一例であると言えます。
図7:トクノシマカンアオイ由来のSBP1とSBP2の3アミノ酸座位を入れ替えると酵素の働きが逆転する。図右側は酢酸鉛試験紙によるMTOXによる生成物(硫化水素)の検出結果。
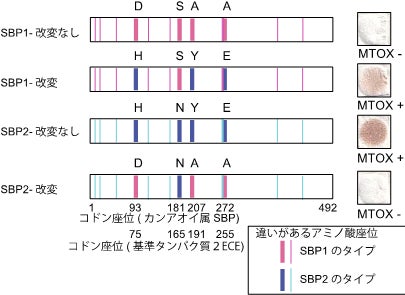
図8:トクノシマカンアオイ、ヒサカキ、カンアオイそれぞれのSBPサブタイプの2ないし3アミノ酸を入れ替えた実験結果。この入れ替えだけでMTOXの働きは失われDSSの働きを獲得することが分かる(それぞれの条件で2回ずつ実験した結果を図示)。
【今後の展望】
本研究では、腐った肉のようなにおいの成分ジメチルジスルフィドを花が放つメカニズムを解明しましたが、そもそも腐った肉がジメチルジスルフィドを出す仕組み、すなわちバクテリアが動物性タンパク質を分解してジメチルジスルフィドを生み出すメカニズムについてもよく分かっていません。またショクダイオオコンニャクなどのように、SBPが関与していないと考えられるジメチルジスルフィド生合成のメカニズムも存在するようです。今後ジメチルジスルフィドを放つさまざまな生物について比較解析することで、生物が「臭いにおい」を生み出すメカニズムの全貌を明らかにしていきたいと考えています。
また、花ではないものに擬態して花粉を運ぶ昆虫をだます花には、腐肉擬態花の他に、キノコのにおいを出す花や発酵した果実のにおいを出す花、昆虫のメスのフェロモンを出す花など擬態にもさまざまなパターンが存在し、それぞれが「ふつうの花」にはない特殊な香り成分を放つことが知られています。今後、これらについても同様の研究手法を用いることで、重点的に研究されている「モデル植物」の研究ではアプローチが難しい特殊な生命現象の数々を解明していきたいと考えています。
本研究は、日本列島の豊かな生物多様性や、植物園等における栽培植物、飼育・培養生物の戦略的なコレクションが未知の生命現象を解き明かすために決定的に重要な役割を果たしうることを示したものと考えています。今後も我が国の豊かな生物多様性がさまざまな科学的発見の源泉として機能し続けるためにも、社会とつながりながら生物多様性の価値を保全、発見し、また普及する筑波実験植物園のミッションを推し進めていきたいと考えています。
【用語解説・その他】
※1 ジメチルジスルフィド(DMDS)
腐った肉や肉食動物の糞などの臭いにおいの主成分の一つで、硫黄を含む分子。バクテリアが動物性タンパク質を分解する際に多く発生する。一般的にはヒトの鼻には悪臭として捉えられるが、チーズや食肉にもわずかに含まれており微量であれば逆に食欲を刺激することが知られている。ハエが強く誘引されるにおい物質でもあり、ハエに受粉されるショクダイオオコンニャクやラフレシアなどの花からも多く放たれる。
※2 カンアオイ属
カンアオイ属はウマノスズクサ科の植物の一群で、葉の下で、地面すれすれに花を咲かせるのが特徴。この花はまるで世界最大の花として知られるラフレシアのミニチュアのようで、実際にラフレシアの花と同じように臭いにおいを放つものがある。
カンアオイの名前は「寒葵」に由来し、これは徳川家の家紋のモチーフとなっている葵(フタバアオイ)に近縁で、冬でも葉が残る常緑性であることにちなむ。

※3 トランスクリプトーム(全発現遺伝子)解析
ある生物のある組織で働いている(発現している)遺伝子は、メッセンジャーRNA(mRNA)の形で組織に存在し、強く発現している遺伝子はそれだけ多くのmRNA分子として存在する。ある組織から全てのmRNAを抽出し、これを逆転写してDNAの形(相補DNA)にして超並列DNAシーケンサーで網羅的に解読することで、ほぼ全ての遺伝子についてそれぞれどのくらい強く発現しているか、あるいは発現していないかを調べることができる。これをトランスクリプトーム解析と呼ぶ。
※4 遺伝子クローニングと組換えタンパク質
機能を調べたいある生物の特定の遺伝子について、タンパク質をコードしている開始コドンから終止コドンまでの配列をPCR法によって取得し、大腸菌の中で増幅可能なDNA分子であるプラスミドに組み込み、このプラスミドを精製することを遺伝子クローニングと呼ぶ。このプラスミドをタンパク質発現用大腸菌に組み込むことで、さまざまな生物の遺伝子産物を組換えタンパク質という形で大腸菌に大量に作らせることができる。これによって遺伝子産物の酵素としての機能を調べることが可能になる。
※遺伝子と遺伝子産物の表記について
遺伝子名はイタリックで、遺伝子産物(タンパク質)名は立体で表記する。例えば、セレン結合タンパク質遺伝子はSBPと表記し、その遺伝子産物であるセレン結合タンパク質はSBPと表記する。
このプレスリリースには、メディア関係者向けの情報があります
メディアユーザー登録を行うと、企業担当者の連絡先や、イベント・記者会見の情報など様々な特記情報を閲覧できます。※内容はプレスリリースにより異なります。
すべての画像
